http://www.biologydiscussion.com/ecosystem/energy-flow-in-an-ecosystem-with-diagram/6740
Energy has been defined as the capacity to do work. Energy exists in two forms potential and kinetic.
Potential energy is the energy at rest {i.e., stored energy) capable of performing work. Kinetic energy is the energy of motion (free energy).
It results in work performance at the expense of potential energy. Conversion of potential energy into kinetic energy involves the imparting of motion.
The source of energy required by all living organisms is the chemical energy of their food. The chemical energy is obtained by the conversion of the radiant energy of sun.
The radiant energy is in the form of electromagnetic waves which are released from the sun during the transmutation of hydrogen to helium. The chemical energy stored in the food of living organisms is converted into potential energy by the arrangement of the constituent atoms of food in a particular manner. In any ecosystem there should be unidirectional flow of energy.
This energy flow is based on two important Laws of Thermodynamics which are as follows:
(1) The first law of Thermodynamics:
It states that the amount of energy in the universe is constant. It may change from one form to another, but it can neither be created nor destroyed. Light energy can be neither created nor destroyed as it passes through the atmosphere. It may, however, be transformed into another type of energy, such as chemical energy or heat energy. These forms of energy cannot be transformed into electromagnetic radiation.
(2) The second law of Thermodynamics:
It states that non-random energy (mechanical, chemical, radiant energy) cannot be changed without some degradation into heat energy. The change of energy from one form to another takes place in such a way that a part of energy assumes waste form (heat energy). In this way, after transformation the capacity of energy to perform work is decreased. Thus, energy flows from higher to lower level.
Main source of energy is sun. Approximately 57% of sun energy is absorbed in the atmosphere and scattered in the space. Some 35% is spent to heat water and land areas and to evaporate water. Of the approximately 8% of light energy striking plant surface, 10% to 15% is reflected, 5% is transmitted and 80 to 85% is absorbed; and an average of only 2% (0.5 to 3.5%) of the total light energy striking on a leaf is used in photosynthesis and rest is transformed into heat energy.
Energy flow in Ecosystems:
Living organisms can use energy in two forms radiant and fixed energy. Radiant energy is in the form of electromagnetic waves, such as light. Fixed energy is potential chemical energy bound in various organic substances which can be broken down in order to release their energy content.
Organisms that can fix radiant energy utilizing inorganic substances to produce organic molecules are called autotrophs. Organisms that cannot obtain energy from abiotic source but depend on energy-rich organic molecules synthesized by autotrophs are called heterotrophs. Those which obtain energy from living organisms are called consumers and those which obtain energy from dead organisms are called decomposers (Fig. 3.7).
When the light energy falls on the green surfaces of plants, a part of it is transformed into chemical energy which is stored in various organic products in the plants. When the herbivores consume plants as food and convert chemical energy accumulated in plant products into kinetic energy, degradation of energy will occur through its conversion into heat. When herbivores are consumed by carnivores of the first order (secondary consumers) further degradation will occur. Similarly, when primary carnivores are consumed by top carnivores, again energy will be degraded.
Trophic level:
The producers and consumers in ecosystem can be arranged into several feeding groups, each known as trophic level (feeding level). In any ecosystem, producers represent the first trophic level, herbivores present the second trophic level, primary carnivores represent the third trophic level and top carnivores represent the last level.
Food Chain:
In the ecosystem, green plants alone are able to trap in solar energy and convert it into chemical energy. The chemical energy is locked up in the various organic compounds, such as carbohydrates, fats and proteins, present in the green plants. Since virtually all other living organisms depend upon green plants for their energy, the efficiency of plants in any given area in capturing solar energy sets the upper limit to long-term energy flow and biological activity in the community.
The food manufactured by the green plants is utilized by themselves and also by herbivores. Animals feed repeatedly. Herbivores fall prey to some carnivorous animals. In this way one form of life supports the other form. Thus, food from one trophic level reaches to the other trophic level and in this way a chain is established. This is known as food chain.
A food chain may be defined as the transfer of energy and nutrients through a succession of organisms through repeated process of eating and being eaten. In food chain initial link is a green plant or producer which produces chemical energy available to consumers. For example, marsh grass is consumed by grasshopper, the grasshopper is consumed by a bird and that bird is consumed by hawk.
Thus, a food chain is formed which can be written as follows:
Marsh grass → grasshopper → bird → hawk
Food chain in any ecosystem runs directly in which green plants are eaten by herbivores, herbivores are eaten by carnivores and carnivores are eaten by top carnivores. Man forms the terrestrial links of many food chains.
Food chains are of three types:
- Grazing food chain
- Parasitic food chain
- Saprophytic or detritus food chain
- Grazing food chain:
The grazing food chain starts from green plants and from autotrophs it goes to herbivores (primary consumers) to primary carnivores (secondary consumers) and then to secondary carnivores (tertiary consumers) and so on. The gross production of a green plant in an ecosystem may meet three fates—it may be oxidized in respiration, it may be eaten by herbivorous animals and after the death and decay of producers it may be utilized by decomposers and converters and finally released into the environment. In herbivores the assimilated food can be stored as carbohydrates, proteins and fats, and transformed into much more complex organic molecules.
The energy for these transformations is supplied through respiration. As in autotrophs, the energy in herbivores also meets three routes respiration, decay of organic matter by microbes and consumption by the carnivores. Likewise, when the secondary carnivores or tertiary consumers eat primary carnivores, the total energy assimilated by primary carnivores or gross tertiary production follows the same course and its disposition into respiration, decay and further consumption by other carnivores is entirely similar to that of herbivores.
Thus, it is obvious that much of the energy flow in the grazing food chain can be described in terms of trophic levels as outlined below:
 A schematic representation of grazing food chain showing input and losses of energy has been presented in Fig. 3.8.
A schematic representation of grazing food chain showing input and losses of energy has been presented in Fig. 3.8.
- Parasitic food chain:
It goes from large organisms to smaller ones without outright killing as in the case of predator.
- Detritus food chain:
The dead organic remains including metabolic wastes and exudates derived from grazing food chain are generally termed detritus. The energy contained in detritus is not lost in ecosystem as a whole, rather it serves as a source of energy for a group of organisms called detritivores that are separate from the grazing food chain. The food chain so formed is called detritus food chain (Fig. 3.9).
 In some ecosystems more energy flows through the detritus food chain than through grazing food chain. In detritus food chain the energy flow remains as a continuous passage rather than as a stepwise flow between discrete entities. The organisms in the detritus food chain are many and include algae, fungi, bacteria, slime moulds, actinomycetes, protozoa, etc. Detritus organisms ingest pieces of partially decomposed organic matter, digest them partially and after extracting some of the chemical energy in the food to run their metabolism, excrete the remainder in the form of simpler organic molecules.
In some ecosystems more energy flows through the detritus food chain than through grazing food chain. In detritus food chain the energy flow remains as a continuous passage rather than as a stepwise flow between discrete entities. The organisms in the detritus food chain are many and include algae, fungi, bacteria, slime moulds, actinomycetes, protozoa, etc. Detritus organisms ingest pieces of partially decomposed organic matter, digest them partially and after extracting some of the chemical energy in the food to run their metabolism, excrete the remainder in the form of simpler organic molecules.
The waste from one organism can be immediately utilized by a second one which repeats the process. Gradually, the complex organic molecules present in the organic wastes or dead tissues are broken down to much simpler compounds, sometimes to carbon dioxide and water and all that are left are humus. In a normal environment the humus is quite stable and forms an essential part of the soil. Schematic representation of detritus food chain is given in Fig. 3.9.
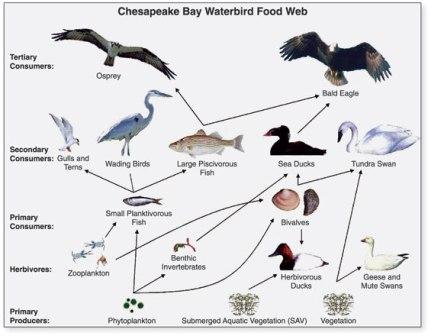
Food web:
Many food chains exist in an ecosystem, but as a matter of fact these food chains are not independent. In ecosystem, one organism does not depend wholly on another. The resources are shared specially at the beginning of the chain. The marsh plants are eaten by variety of insects, birds, mammals and fishes and some of the animals are eaten by several predators.
 Similarly, in the food chain grass → mouse → snakes → owls, sometimes mice are not eaten by snakes but directly by owls. This type of interrelationship interlinks the individuals of the whole community. In this way, food chains become interlinked. A complex of interrelated food chains makes up a food web. Food web maintains the stability of the ecosystem. The greater the number of alternative pathways the more stable is the community of living things. Fig. 3.10. illustrates a food web in ecosystem.
Similarly, in the food chain grass → mouse → snakes → owls, sometimes mice are not eaten by snakes but directly by owls. This type of interrelationship interlinks the individuals of the whole community. In this way, food chains become interlinked. A complex of interrelated food chains makes up a food web. Food web maintains the stability of the ecosystem. The greater the number of alternative pathways the more stable is the community of living things. Fig. 3.10. illustrates a food web in ecosystem.
Ecological pyramid:
The trophic structure of an ecosystem can be indicated by means of ecological pyramid. At each step in the food chain a considerable fraction of the potential energy is lost as heat. As a result, organisms in each trophic level pass on lesser energy to the next trophic level than they actually receive. This limits the number of steps in any food chain to 4 or 5. Longer the food chain the lesser energy is available for final members. Because of this tapering off of available energy in the food chain a pyramid is formed that is known as ecological pyramid. The higher the steps in the ecological pyramid the lower will be the number of individuals and the larger their size.
The idea of ecological pyramids was advanced by C.E. Elton (1927). There are different types of ecological pyramids. In each ecological pyramid, producer level forms the base and successive levels make up the apex. Three types of pyramidal relations may be found among the organisms at different levels in the ecosystem.
These are as follows:
- Pyramid of numbers,
- Pyramid of biomass (biomass is the weight of living organisms), and
- Pyramid of energy.
- Pyramid of numbers:
 It depicts the numbers of individuals in producers and in different orders of consumers in an ecosystem. The base of pyramid is represented by producers which are the most abundant. In the successive levels of consumers, the number of organisms goes on decreasing rapidly until there are a few carnivores.
It depicts the numbers of individuals in producers and in different orders of consumers in an ecosystem. The base of pyramid is represented by producers which are the most abundant. In the successive levels of consumers, the number of organisms goes on decreasing rapidly until there are a few carnivores.
The pyramid of numbers of an ecosystem indicates that the producers are ingested in large numbers by smaller numbers of primary consumers. These primary consumers are eaten by relatively smaller number of secondary consumers and these secondary consumers, in turn, are consumed by only a few tertiary consumers (Fig. 3.11, 3.12a).
This type of pyramid is best presented by taking an example of Lake Ecosystem. In this type of pyramid the base trophic level is occupied by producer elements—algae, diatoms and other hydrophytes which are most abundant. At the second trophic level come the herbivores or zooplanktons which are lesser in number than producers.
 The third trophic level is occupied by carnivores which are still smaller in number than the herbivores and the top is occupied by a few top carnivores. Thus, in the ecological pyramid of numbers there is a relative reduction in number of organisms and an increase in the size of body from base to apex of the pyramid. In parasitic food chain starting from tree, the pyramid of numbers will be inverted (Fig. 3.12).
The third trophic level is occupied by carnivores which are still smaller in number than the herbivores and the top is occupied by a few top carnivores. Thus, in the ecological pyramid of numbers there is a relative reduction in number of organisms and an increase in the size of body from base to apex of the pyramid. In parasitic food chain starting from tree, the pyramid of numbers will be inverted (Fig. 3.12).
- Pyramid of biomass of organisms:
 The living weights or biomass of the members of the food chain present at any one time form the pyramid of biomass of organisms. This indicates, by weight or other means of measuring materials, the total bulk of organisms or fixed energy present at one time. Pyramid of biomass indicates the decrease of biomass in each tropic level from base to apex, e.g., total biomass of producers is more than the total biomass of the herbivores.
The living weights or biomass of the members of the food chain present at any one time form the pyramid of biomass of organisms. This indicates, by weight or other means of measuring materials, the total bulk of organisms or fixed energy present at one time. Pyramid of biomass indicates the decrease of biomass in each tropic level from base to apex, e.g., total biomass of producers is more than the total biomass of the herbivores.
Likewise, the total biomass of secondary consumers will be lesser than that of herbivores and so on (Fig. 3.13, 3.14 a, b). Since some energy and material are lost in each successive link, the total mass supported at each level is limited by the rate at which the energy is being stored below. This usually gives sloping pyramid for most of the communities in terrestrial and shallow water ecosystems. The pyramid of biomass in a pond ecosystem will be inverted as shown in Fig. 3.13 b.
- Pyramid of energy:
This depicts not only the amount of total energy utilized by the organisms at each trophic level of food chain but more important, the actual role of various organisms in transfer of energy. At the producer level the total energy will be much greater than the energy at the successive higher trophic level.
 Some producer organisms may have small biomass but the total energy they assimilate and pass on to consumers may be greater than that of organisms with much larger biomass. Higher trophic levels are more efficient in energy utilization but much heat is lost in energy transfer. Energy loss by respiration also progressively increases from lower to higher trophic states (Fig. 3.15).
Some producer organisms may have small biomass but the total energy they assimilate and pass on to consumers may be greater than that of organisms with much larger biomass. Higher trophic levels are more efficient in energy utilization but much heat is lost in energy transfer. Energy loss by respiration also progressively increases from lower to higher trophic states (Fig. 3.15).
In the energy flow process, two things become obvious. Firstly there is one way along which energy moves i.e. unidirectional flow of energy. Energy comes in the ecosystem from outside source i.e. sun. The energy captured by autotrophs does not go back to the sun, the energy that passes from autotrophs to herbivores does not revert back and as it moves progressively through the various trophic levels, it is no longer available to the previous levels.
Thus due to unidirectional flow of energy, the system would collapse if the supply from primary source, the sun is cut off. Secondly, there occurs a progressive decrease in energy level at each trophic level which is accounted largely by the energy dissipated as heat in metabolic activities.
Productivity:
The relationship between the amount of energy accumulated and the amount of energy utilized within one trophic level of food chain has an important bearing on how much energy from one trophic level passes on to the next trophic level in the food chain. The ratio of output of energy to input of energy is referred to as ecological efficiency.
Different kinds of efficiencies can be measured by the following parameters:
(i) Ingestion which indicates the quantity of food or energy taken by trophic level. This is also called exploitation efficiency.
(ii) Assimilation indicates the amount of food absorbed and fixed into energy rich organic substances which are stored or combined with other molecules to build complex molecules such as proteins, fats etc.
(iii) Respiration which indicates the energy lost in metabolism.
Primary Productivity:
The fraction of fixed energy a trophic level passes on to the next trophic level is called production. Green plants fix solar energy and accumulate it in organic forms as chemical energy. Since it is the first and basic form of energy storage, the rate at which the energy accumulates in the green plants or producers is known as primary productivity.
Primary productivity is the rate at which energy is bound or organic material is created by photosynthesis per unit area of earth’s surface per unit time. It is most often expressed as energy in calories / cm2 / yr or dry organic matter in g / m2 / yr (g/m2 x 8.92 = lb / acre). The amount of organic matter present at a given time per unit area is called standing crop or biomass and as such productivity, which is a rate, is quite different from biomass or standing crop.
The standing crop is usually expressed as dry weight in g/m2 or kg/m2or t/ha (metric tons) or 106g/hectare. Primary productivity is the result of photosynthesis by green plants including algae of different colours. Bacterial photosynthesis or chemosynthesis, although of small significance may also contribute to primary productivity. The total solar energy trapped in the food material by photosynthesis is referred to as gross primary productivity (G.P.P.).
A good fraction of gross primary production is utilized in respiration of green plants. The amount of energy bound in organic matter per unit area and time that is left after respiration in plants is net primary production (N.P.P.) or plant growth. Only the net primary productivity is available for harvest by man and other animals. Net productivity of energy = gross productivity—energy lost in respiration.
Secondary Productivity:
The rates at which the heterotrophic organisms resynthesize the energy-yielding substances is termed as secondary productivity. Secondary productivities are the productivities of animals and saprobes in communities. The amount of energy stored in the tissues of consumers or heterotrophs is termed as net secondary production and the total plant material ingested by herbivores is grass secondary production. Total plant material ingested by herbivores minus the materials lost as faeces is equal to Ingested Secondary Production.
Environmental factors affecting the production processes in an ecosystem are as follows:
- Solar radiation and Temperature
- Moisture. Leaf water potential, soil moisture and precipitation fluctuation and transpiration.
- Mineral nutrition. Uptake of minerals from the soil, rhizosphere effects, fire effects, salinity, heavy metals, nitrogen metabolism.
- Biotic activities. Grazing, above ground herbivores, below ground herbivores, predators and parasites, diseases of primary producers.
- Impact of human population. Pollutions of different sorts, ionizing radiations like atomic explosions, etc.
There are three fundamental concepts of productivity:
- Standing crop
- Materials removed
- Production rate.
- Standing crop:
It is the abundance of the organisms existing in the area at any one time. It may be expressed in terms of number of individuals, as biomass of organisms, as energy content or in some other suitable terms. Measurement of standing crop reveals the concentration of individuals in the various populations of ecosystem.
- The materials removed:
The second concept of productivity is the materials removed from the area per unit time. It includes the yield to man, organisms removed from the ecosystem by migration, and the material withdrawn as organic deposit.
- The production rate:
The third concept of productivity is the production rate. It is the rate at which the growth processes are going forward within the area. The amount of material formed by each link in the food chain per unit of time per unit area or volume is the production rate.
All the three major groups of organisms—producers, consumers and reducers are the functional kingdoms of natural communities. The three represent major directions of evolution and are characterised by different modes of nutrition. Plants feed primarily by photosynthesis, animals feed primarily by ingesting food that is digested and absorbed in the alimentary canal and the saprobes feed by absorption and have need for an extensive surface of absorption. The principal kinds of organisms among saprobes are the unicellular bacteria, yeasts, chytrids or lower fungi and higher fungi with mycelial bodies.
In terrestrial communities as much as 90% of net primary production remains un-harvested and are utilized as dead tissue by saprobes and soil animals. The saprobes have a larger and more essential role than animals in degrading dead organic matter to inorganic forms and in such ecosystems, secondary production by reducers (decomposers) should exceed that by consumers, though the former is even more difficult to measure than the latter.
Biomass of decomposers with their microscopic cells and filaments embedded in food sources is also difficult to measure and that is small in relation to their productivity and significance for the ecosystem. Small masses of reducers degrade and transform larger masses of organic matter to inorganic remnants. In so doing decomposers disperse back to the environment the energy of photosynthesis accumulated in the organic compounds that are decomposed.
Thus they have a major role in the energy flow of ecosystems. A community or ecosystem, like an organism, is an open energy system. The continuous intake of energy in photosynthesis replaces the energy dissipated to environment by respiration and biological activity and the system does not run-down through the loss of free energy to maximum entropy.
If the amount of energy entrapped is greater than the energy dissipated, the pool of biologically useful energy of organic bonds increases. This results in increase of community biomass and consequently the community grows; such is the case in succession. If energy intake is lesser than energy dissipation, the community biomass will decrease and it must, in some sense, retrogress. If energy intake and loss are in balance, the pool of organic energy is in steady state; such is the case in climax communities.
Three aspects of this steady state may be recognized:
(i) The steady state of population of climax communities in which equal birth and death rates in population keep the number of individuals relatively constant,
(ii) The steady state of energy flow,
(iii) The steady state of the matter of community, where addition of material by photosynthesis and organic synthesis is balanced by loss of material through respiration and decomposition.
Methods of Measuring Primary Production:
There are several parameters for measuring primary production and the methods of measuring primary production are based on those parameters.
The methods are discussed here as under:
- Harvest method:
It involves removal of vegetation periodically and weighing the material. For measuring above ground production, the above ground plant parts are clipped at ground level, dried to constant weight at 80°C and weighed. The dry weight in g/m2 /year gives the ground production. Below ground production is estimated by using frequent core sampling technique of Dahlman and Kucera (1965). It is expressed in terms of weight in gm per unit area per year. In terms of energy one gm dry weight of plant material contains 4 to 5 kcal.
The limitations of harvest method are as follows:
(i) The amount of plant material consumed by herbivores and the food oxidized during respiration process of the plants is not accounted.
(ii) Root biomass is neglected.
(iii) Photosynthetic trans located to underground parts of plants are not known.
In spite of these limitations the method is used all over for measuring net assimilation rate (NAR) and relative growth rate (RGR).
- Carbon dioxide assimilation method:
Utilization of CO2 in photosynthesis or its liberation during respiration is measured by infrared gas analysis or by passing the gas through Baryta water Ba(OH)2 and titrating the same. The CO2removed from incoming gas chamber is taken to be synthesized into organic matter by the green plants. Performing the experiment in light and dark chambers the net and gross production can be measured.
In the lighted chamber photosynthesis and respiration take place simultaneously and the CO2 coming out from the chamber is the unused gas of the atmosphere plus gas from the respiration of plant parts. In the dark chamber all CO2 is due to respiration.
Net production = Gross production—Respiration
- Oxygen production method:
In the aquatic vegetation CO2 gas analysis method is not used but oxygen evolution method is generally used. The light and dark bottle technique is employed for measuring primary production of aquatic plant. In this method two bottles, one transparent and the other opaque are filled with water at a given depth of lake, closed, maintained at that depth for some time and then brought to laboratory for determination of oxygen content in the water. The decrease of oxygen in dark bottle is due to respiratory activity while increase of O2 in light bottle is due to photosynthesis. The total increase of O2 in light bottle plus the amount of O2 decreased in dark bottle express gross productivity (O2 value multiplied by 0.375 gives an equivalent of carbon assimilation). Recently, oxygen electrodes have been used for estimating oxygen content in water.
- Chlorophyll method:
Gesner (1949) pointed out that the amount of chlorophyll/m2 is almost limited to a narrow range of 0.1 to 3.0 gm regardless of the age of individuals or the species present therein. There is direct correlation between the amount of chlorophyll and dry matter production in different types of communities with varying light conditions.
The relation of total amount of chlorophyll to the photosynthetic rate is referred to as assimilation ratio or rate of production/gm chlorophyll. Total chlorophyll per unit area is greater in land plants as compared to that in aquatic plants. In marine ecosystem the rate of carbon assimilation is 3.7 g/ hr/g of chlorophyll. The relationship between area based chlorophyll and dry matter production in terrestrial ecosystems has been worked out by Japanese ecologists Argua and Monsi (1963).
- Other methods:
Pandeya (1971), Sharma (1972) and several other ecologists have evolved correlation coefficients for evaluating biomass and productivity in forest trees by measuring their diameter at breast height (DBH), height, canopy cover, etc.
Methods of establishing regression are as below:
(i) Diameter of trees in sample quadrats is measured at breast height and the height repeated is determined for each tree.
(ii) Different diameter and height classes are determined for each species.
(iii) A set of sample trees are cut and subjected to a detailed analysis for dry weight of stems, twigs, leaves and roots.
(iv) Regression values are computed for the sets of trees belonging to each girth class, relating the biomass of each fraction to the diameter at breast height.
(v) The regression values are used to compute the probable biomass and Production each tree in the sample area. These values for each species when pooled give biomass and production rate of trees per unit area in the forest. Age of the trees markedly influences the annual net production.



